
The Genus Opuntia
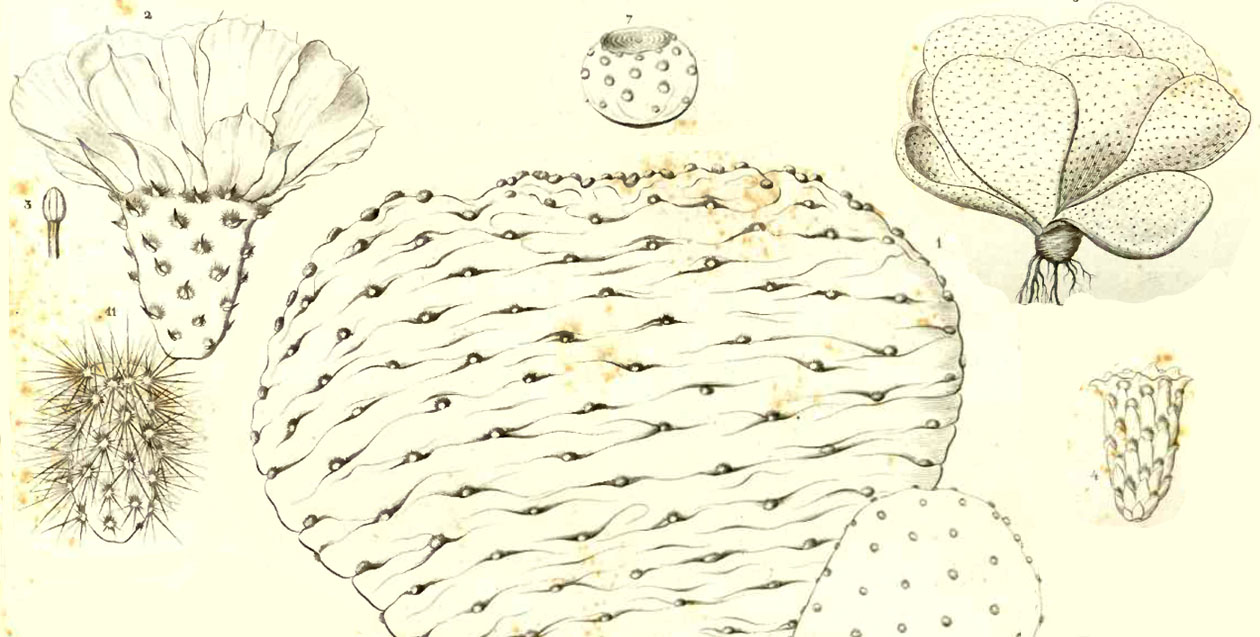
Modified from Engelmann & Bigelow 1856
Opuntia basilaris Engelmann & Bigelow
Mescaline is present in trace amounts.

Opuntia basilaris
Common name: “Beavertail cactus”
Etymology: In reference to the branching arising from the base.
Distribution & occurrence: Arizona, California, Nevada and Utah. Also in Sonora, Mexico. Entry #213 in Piazzetti 1985.
Occurs in southwestern US and into northern Mexico (Sonora).

Opuntia basilaris
” humilis, articulis obovatis s. subtriangularibus glaucescentibus pubescentibus adscendentibus e basi proliferis, fere rosulatis ; foliis subulatis minutis erectis rubellis tomentum axillare vix superantibus ; pulvillis subconfertis fulvo-tomentosis setas gracillimas demum numerosissimas breves fulvidas et subinde aculeolos setiformes caducos gerentibus ; floris purpurei ; ovario obovato pubescente pulvillis plurimis (40-60) confertis fulvtomentosis instructo, sepalis 20-25 exterioribus oblanceolatis acuminatis, interioribus late obovatis cuspidatis, petalis sub-10 obov-orbiculatis retusis s. obcordatis saepe tenuiter mucronatis, stigmatibus 8 brevibus in capitulum conicum congestis ; bacca (sicca?) breviter obovata late umbilicata, seminibus magnis crassis subregularibus. (Plate XIII, fig. 1-5 & XXIII fig. 14)
On hills and in ravines from the Cactus Pass down the valley of Williams river to the Colorado, and to Mojave creek ; Mr. Schott met with it on the lower Gila ; and both he and Mr. Albert H. Campbell obtained the beautiful purple flowers of this plant in April and May, 1855. The habit of this plant is very different from any other of our Opuntiae, as the stout obovate or often fan-shaped or sometimes almost obcordate joints originate from a common base form a kind of rosette, resembling somewhat an open cabbage head. […] Joints 5-8 inches long, ½ inch in thickness, minutely pubescent ; leaves only 1 line in length, slenderly subulate, smaller than any other of our species […].
Pulvilli somewhat immersed, 4-6 lines apart. Flower of a beautiful and rich purple color, about 2½ inches in diameter, ovary nearly 1 inch long, crowded with 40-60 elevated areolae, with light brown wool and brighter brown bristles ; filaments not very numerous, leaving the inner base of the tube naked ; stigmata about 2 lines long, or less, apparently green. Fruit seems to be perfectly dry, short and thick; seeds 3 lines in diameter, nearly 2 lines thick, with a rather narrow but very thick rim, regular or sometimes quite irregular.” pp. 43-44, Engelmann & Bigelow 1856. (“Lines” refer to increments of o.1″.)

Opuntia basilaris
“c. : humilis ; articulis obovatis seu triangularibus glaucescentibus pubescentibus e basi proliferis; foliis minutis ; puvillis subconfertis fulvo-villosis setas gracillimas demum numerosissimas fulvidas et subinde aculeolos setiformes caducos gerentibus ; floris purpurei ovario obovato pulvillis plurimis instructo; stigmatibus 8 in capitulum congestis; bacca obovata late umbilicata (sicca?) ; seminibus magnis crassis subregularibus.
0n Williams’s River, the Colorado, and the Mojave, and down to the Gila : fl. April and May. – Habit very different from any other of our Opuntiae ; the stout obovate or fan-shaped joints (5 – 8 inches long) originate from a common base, forming a sort of rosette.Leaves only one line long, 4 – 6 lines apart ; pulvilli red-brown, somewhat immersed. Flower about 2½ inches in diameter; ovary with 40 – 60 pulvilli. Fruit apparently dry, thereby approaching the next section. Seed 3 lines in diameter, 2 lines thick.” pp. 298-299, Engelmann & Bigelow 1856 PAAS.

Opuntia basilaris
Very striking low growing plants. Usually grayish to bluish green, often reddened around areoles. Almost appearing spineless from a distance due to the clumps of short glochids and laterally wrinkled skin of the pads. A number of varieties exist with varying degrees of spination (some having only short lived glochids and no spines), pad forms, flowers and fruit.
Cerise flowers are most common. Other colors are known in some varieties. Flowering time variable. Usually after it gets warm. Flowers are usually 5 cm in diameter. Fruit is short and oval shaped. Large and thick seeds.
Several distinct varieties are recognized.

Opuntia basilaris
W. Hubert Earle 1980 and Lyman Benson 1982 and Curt Backeberg 1977; Schuster 1990, page 146, has a photograph of Opuntia basilaris in flower; in habitat next to Opuntia imbricata.
At least one variety, (var. cordata) is a horticultural cultivar.
Stays small and is fairly cold tolerant. Pizzetti 1985.

Opuntia basilaris
Did you catch the divergence of those plants from what was said in the original description?
Reported analysis
tlc did not indicate the presence of any alkaloids.
Tandem mass spectrometry detected mescaline at 0.01% by dry weight and
3,5-Dimethoxyphenethylamine at less than 0.01% dry weight.
Ma et al. 1986
It was not noted by Ma and coworkers which variety they analyzed.
-

- Opuntia basilaris
-

- Opuntia basilaris
-

- Opuntia basilaris
Opuntia ficus-indica (Linnaeus) Miller
Philip Miller (1768) Gardeners Dictionary, Edition 8, no. 2, as Opuntia ficus-indica.
Caroli Linnaei (1751) Species Plantarum, 1: 468, as Cactus ficus indica.
The presence of mescaline was reported.

Opuntia ficus-indica
Common names: “Mission cactus” “Tuna” “Barbary Fig” , “Indian Fig” “Prickly Pear” (US) [Many of the names are also applied to the fruit] “Barshoom” (Saudi Arabia)
Etymology: ficus-indica means “Indian fig” in reference to the edible fruit.
Distribution & occurrence: Origin unknown.
Believed to be tropical America (it is clear that it underwent intensive human development in Mexico). Cultivated widely and occasionally naturalized in the Mediterranean and temperate to tropical regions.
Has widely and commonly (and readily) hybridized with native
Opuntias in southern California but, in US, only common as an escaped plant in Hawaii.
Considered a weedy pest in Australia and parts of Africa where biological warfare in the form of deliberate cochineal and Cactoblastis releases have been employed.

Opuntia ficus-indica
“articulis ovato-oblongis, spinis setaceis. Indian Fig with oblong oval joints, and bristly spines. Opuntia folio oblongo media. Tourn. Inst. R.H. 239. Middle Indian Fig with oblong leaves.”
Miller 1753.
” Cactus articulato-prolifer ; articulis ovato-oblongis, spinis setaceis.
Cactus compressus articulatis ramosus, articulis ovato-oblongis : spinis setaceis Hort. cliff. 183. Hort ups. 120 Roy lugdh. 280.
Habitat in America, calidiora. [saturn symbol]”
p. 468, Linnaei 1751.

Opuntia ficus-indica
Shrubby plants, 3-5 ft., that may become tree-like (to 16.5 feet: Pizzetti; to 7 meters: Benson)
Trunk is woody up to 120 cm long and 120 cm in diameter, and much branched on upper part.
Oval or oblong thick and fleshy pads can reach 16 inches.
Small areoles normally lack spines but possess many yellow glochids which fall off with age.
A spiny type is also known to exist. It has 1-6 white spines [Note
39] per areole that can reach 4 cm but are usually shorter. Shorter spined plants are believed to have developed by Mexican horticulturalists.
Yellow to yellow-orange flowers are large; with pale yellow stamens.
Fruit is “pear-shaped and umbillicate at its apex”. It can be yellow, [pale yellow-green (de Castilla variety)], orange, red, reddish, purplish or striped. The pulp is normally the same color as the epidermis. [Ed.: When striped?]
Widely cultivated. Benson says “prized for their fruit since prehistoric times.” Claimed the best edible cactus fruit [ed.: this might be disputed].
Multiple cultivars exist; some are grown for fruit and some for their edible pads.
“Burbank’s Thornless” is a common and completely thornless type that is popular for food and landscaping. The war that was waged on Burbank by the USDA (and William Safford) over this particular selection bears mention but is outside of the scope of this entry.
[Flath & Takahashi 1976 note the flavor of the de Castilla variety to be mildly sweet and melon-like with little acid character.
They describe its flesh as pale-green and similar in texture to a ripe melon or kiwi. Interestingly they found it to share some volatile C-9 compounds which are also known to occur in cucumbers and certain melons and which are claimed responsible for the character of their flavor.]
A more detailed look at this plant can be found in Cactus Chemistry By Species.

Opuntia ficus-indica
Benson notes that it requires deep soil and some subirrigation in order for it to escape and become a problem. Its entry into the gene pool of the hybrid swarms of southern California was due to the ease of insect cross pollination with other, already weedy, Opuntia species.
Benson 1982: Entry #46, pages 512-516, hybrids, 517-528. Detailed description and heavily illustrated with pictures. Color plates of spiny and spineless forms, color plates 56 and 54 respectively. Fruit: 55; Species with native and hybrid swarm: 57
and Pizzetti 1985: Entry # 219 (Includes picture)
Available varieties (offered by Koehres) include:
v. alba
v. rubra
v. sanguinea
Cultivation:
Requires strong sun and perfect drainage.
Pizzetti recommends a sandy or stony soil.

Opuntia ficus-indica
Water content:
Pads determined to contain 87.4% (young) and 85.4% (mature) water by weight. Kircher 1982

Opuntia ficus-indica
Published analysis:
Opuntia ficus-indica has been reported to contain:
Mescaline (%?)
N-Methyltyramine (%?)
Tyramine (%?)
El-Moghazy et al. 1982 analyzed Egyptian-grown plants but did not give the percentages recovered.
Lipid content determined to be 2.5% by dry weight: Kircher 1982
Unidentified lactone-forming acid (tlc) Kringstad & Nordal 1975
Isorhamnetin was found in the hydrosylate of flower pigments.
Arcoleo et al. 1961
β-Sitosterol Dawider & Fayez 1961; (0.04% dry wt. in flowers)
Arcoleo 1966
free Lauric, Myristic, Palmitic, Stearic and Oleic acids & also
the esters of Myristic, Palmitic, Stearic, and Oleic acid. Arcoleo 1966
Myrcene, Limonene & γ-Terpinene (terpenes: small amounts in the de Castilla variety fruit) Flath & Takahashi 1978. [They also reported other volatile compounds in the fruit including many alcohols, aldehydes, ketones, esters & hydrocarbons such as Toluene & Methylcyclohexane]
Fruit contains betacyanins: Betanin (major) & Isobetanin. Piattelli & Minale 1964a & 1964b.
Indicaxanthin (a betaxanthin) was reported in mature fruit (orange yellow variety) by Impellizzeri & Piattelli 1972. (Also in Piattelli et al. 1964a & 1964b)
Indicaxanthin & Betanin in fruit. Minale et al. 1965
Malic acid, Citric acid, Piscidic acid, Piscidic acid monoethyl ester (0.0433%), Piscidic acid diethyl ester (0.0333%) and several other nonvolatile acids in fruit. Nordal et al. 1966.
de Castilla fruit showed pH 4.85-6.3.
Citric acid was reported at levels of 0.084-0.12% according to Flath & Takahashi 1976.
Opuntia ficus-indica is widely used for food and is incorporated in many folk remedies (frequently based on the mucilaginous properties of the pulp).
In Bolivia, the pulp is used as a refrigerant and anti-inflammatory for healing intestinal and stomach problems. Parboiled, peeled leaves sprinkled with vinegar or camphorated alcohol is applied as a plaster to treat angina, malignant tumors and kidney pains.
It is said that if the leaf pulp is “abused” it causes diarrhea, lassitude, and weakness in all body parts as well as provoking profound dreams [Note 40].
Salcedo 1986

A thirsty Opuntia ficus-indica in Oz

















































































































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}